|
|
|
Reference: Hackney, A.C. Testosterone and reproductive dysfunction in endurance-trained men. In: Encyclopedia of Sports Medicine and Science, T.D. Fahey (Editor). Internet Society for Sport Science: http://sportsci.org. 20 Sept 1998.
Reviewed by Kevin Guskiewicz, Sports
Medicine, University of North Carolina, Chapel Hill, North Carolina,
USA.
Basic Endocrinology
and Physiology
Methodological
Concerns
Exercise and
Testosterone
Physiological Impact of
Lower Testosterone
Conclusions
Further Reading
Endurance training can result in dysfunction in the reproductive system of humans. Research within the sports medicine community has primarily focused upon reproductive dysfunction associated with athletic women. Not until recently have studies seriously addressed the question of how exercise training affects the male reproductive system. Research findings in this area have led some investigators to suggest that the effect of exercise training on the male reproductive system may be comparable to some degree to that found in women. For example, endurance-trained athletes of both sexes have abnormally low levels of the major sex hormones: testosterone in men, and estrogen in women.
The objective of this paper is to present an overview of how endurance exercise training affects testosterone and other aspects of the male reproductive system. The material is divided into four major sections. The first section deals with the basic endocrinology and physiology of testosterone in the male. The second section deals with physiological and methodological issues surrounding the interpretation of research on hormones. The third section covers the changes in testosterone in response to exercise training and acute or prolonged exercise bouts. Finally, in the fourth section the physiological impact of the changes observed in testosterone levels associated with endurance training are discussed.
Basic Endocrinology and Physiology
Regulation of Testosterone Production
Testosterone is a steroid hormone. Other steroids in the body include cholesterol, bile acids, vitamin D, and hormones of the adrenal glands and ovary. The majority of circulating testosterone in men comes from production in the interstitial cells of Leydig at the testicles. The adrenal gland also produces small amounts. Regulation of testicular production occurs via a negative feedback loop system involving the anterior pituitary, hypothalamus, and testicles; referred to as the hypothalamic-pituitary-testicular axis.
Periodically the hypothalamus releases pulses of gonadotrophin-releasing hormone (GnRH) into the hypophyseal circulation, which supplies the hypothalamus and anterior pituitary. The GnRH stimulates the anterior pituitary to produce and release luteinizing hormone (LH) and follicle-stimulating hormone (FSH). The pulsatile release of GnRH results in LH and FSH also being released into the systemic circulation in a similar pulsatile manner. For normal, healthy males approximately 2 to 4 LH and FSH pulses are observed over a 6- to 8-hour period, however the amplitudes of the LH pulses are much greater than those observed for FSH. At the testicles, LH and FSH interact with their primary target tissue receptors (LH, Leydig cells; FSH, Sertoli cells) located on the respective cell membranes. Once a hormone-receptor complex is formed, there is an adenyl cyclase-mediated increase of cyclic AMP, which produces a phosphorylation of intracellular proteins by activation of a protein kinase mechanism. In the Leydig cells this protein kinase activation leads to a mobilization of steroid precursors, in particular the activation of pregnenolone synthesis from cholesterol. Pregnenolone serves as the parent compound from which testosterone is derived. Several other hormones participate in these regulatory events. One worth noting is the anterior pituitary hormone prolactin, which in low concentrations acts as a potentiator of LH at the Leydig cells. This hormone will be discussed later in this article.
Synthesized testosterone diffuses from the Leydig cells into the testicular vascular system and/or into adjacent testicle compartments containing the Sertoli cells. In the Sertoli cells, testosterone plays an essential role in the facilitation of the spermatogenesis process. (The FSH receptor-hormone formation at the Sertoli cell results in the initiation of the spermatogenesis process.) The pulsatile release of LH results in some fluctuations of testosterone levels in the circulation and additionally there are circadian cycles in which large nocturnal elevations in testosterone can be observed
The majority of the circulating testosterone is transported bound to various carrier proteins (sometimes referred to as binding proteins). The principal carrier protein is sex steroid-binding globulin (SSBG), however other plasma proteins can also bind and carry testosterone to a lesser degree (e.g., albumin, cortisol-binding globulin). The remaining non-bound circulating testosterone is referred to as free testosterone. This free testosterone is considered the biologically active form of the hormone, as this portion of the hormone can interact at the target tissue receptors. Circulating bound and free testosterone are collectively referred to as total testosterone; sometimes in the literature the term testosterone is used synonymously for total testosterone. Table 1 presents a short summary of the common abbreviations used in this area of endocrinology.
|
Table 1: Common abbreviations used in reproductive endocrinology as associated with exercise studies and used within this text. | ||
|
Abbreviation |
Full Name |
Gland/Tissue of Origin |
|
T |
Testosterone |
Testes |
|
LH |
Luteinizing hormone |
Pituitary |
|
FSH |
Follicle stimulating hormone |
Pituitary |
|
GnRH |
Gonadotrophin-releasing hormone |
Hypothalamus |
|
C |
Cortisol |
Adrenal cortex |
|
SSBG (or SHBG) |
Sex steroid-binding globulin (sex hormone-binding globulin) |
Liver |
Metabolic Roles of Testosterone
Testosterone has several physiological roles within the male. These roles can be divided into two major categories: androgenic effects, related to reproductive function and the development of a male's secondary sex characteristics, and anabolic effects, pertaining more generally to stimulation of tissue growth.
Androgenic Effects. A major reproductive role of testosterone involves the development of the sperm cell. At the Sertoli cells of the testicles, testosterone induces a nuclear activation process which stimulates and catalyses the maturation and development of sperm during the process of spermatogenesis. Maintenance of testosterone levels within the Sertoli cells is essential for the development of adequate numbers of mature, viable sperm that are necessary for a male to be fertile.
Testosterone also assists in the development and functioning of the male accessory sex glands (prostate, seminal vesicles, and epididymides), which aid in the sperm development and function, as well as in the act of copulation. Also attributed to the influence of testosterone are the secondary sex characteristics of males, such as the typical deeper male voice, increased levels of body hair, penile growth, sex drive (libido), and more aggressive behavior patterns.
Testosterone also helps red-blood cell development and can produce increased sodium reabsorption in the kidneys. Animal based research has also suggested testosterone plays a role in the increase of tissue glycogen synthesis and storage.
Effectors of Testosterone Levels: Physiological Concerns
The level of testosterone in the circulation is a function of the amount of hormone entering (testicular production and secretion) and the amount leaving (metabolic clearance) the blood pool. The rates for these processes are affected by any changes in the physiological state that alter metabolic turnover of the hormone.
Production and Secretion. Testicular testosterone production in the normal, average male is approximately 7 mg per day. This rate is affected by circulating production stimulators (LH), testicle LH receptor numbers, and synthesis substrate availability.
LH and testosterone levels are directly related. However, it is believed that only a small percentage of the total testicular LH receptors need to be occupied to achieve maximum stimulation of the Leydig cells. Reductions in the numbers of testicular LH receptors can result in reduced levels of testosterone production even in the presence of elevated LH. Receptor numbers can be reduced by certain pathological conditions (e.g. hypergonadotrophic hypogonadism) or a down-receptor phenomena" related state. Persistent elevations in circulating LH can produce a down-regulation of receptor numbers and thereby reduce testicular sensitivity to further changes in blood LH levels. The conditions of hyperprolactinaemic and hypercortisolemic can also produce lowered testosterone levels. The exact mechanism for this lower testosterone effect is uncertain, but may be due to interference with Leydig cell receptors and/or direct inhibition of the steroidogenic synthesis pathway. Finally, any reduction in availability of steroid hormone precursors (cholesterol, and pregnenolone) can produce a lower rate of testosterone synthesis.
Secretion is affected mainly by testicular blood flow through the testicular area, since testosterone is lipid soluble and thus freely diffusible. Furthermore, the testicles apparently have little or no storage capacity for testosterone. Testicular blood flow moreover is a function of the levels of vascular vasoconstriction or vasodilatation. Therefore, anything that influences vascular tone can affect the rate of testosterone secretion.
Metabolic Clearance. Testosterone is cleared from the blood by uptake in target tissues and by degradation in the liver. The degradation process involves the conversion of testosterone into 17-ketosteroids and glucuronide, which are excreted into the urine. The hepatic removal process is primarily a function of hepatic blood flow. Thus any changes in the hepatic blood flow can result in concomitant changes in the removal rate of testosterone. Moreover, a small portion of the testosterone is removed from the circulation by select body tissues and then converted to estradiols.
It is also important to realize changes observed in blood levels of testosterone may not necessarily reflect alterations in production, secretion, or metabolic clearance rates. A prime example, relative to exercise studies, involves the effect of shifts in plasma. Owing to the binding of testosterone to carrier proteins, increases or decreases in plasma volume lead to a dilution or concentration that is not indicative of changes in the normal hormonal turnover rate. Whether these changes in concentration have a physiological impact is a point of debate.
When discussing factors affecting blood levels of testosterone, consideration must be given to non-physiological factors that could be sources of variation between studies. Examples include blood sampling method, diurnal variations in hormone concentrations, hormone detection methodology, and research protocol.
The timing and method of blood sampling are considerations that must be carefully addressed when comparing or evaluating testosterone results of different studies. The pulsatile release of LH produces fluctuations in the circulating levels of testosterone. Furthermore, as noted earlier there is a daily circadian pattern in blood testosterone levels. A single blood sample will therefore provide an imprecise estimate of the average hormonal level. Use of multiple or serial blood sampling will provide a more accurate assessment. Ideally, the use of a serial method of blood sampling would be preferred and considered more informative of the true status of circulating testosterone.
The hormonal biochemical assay methodology must also be considered a source of variability in results. Currently, radioimmunoassays (RIA) are the most common means of assessing testosterone concentrations in blood samples. Since the development of the technique, over 30 years ago, tremendous refinements in procedural and technical aspects have occurred and are still constantly happening. Hence, comparison of hormonal values between studies becomes complicated if a number of years separates the experiments. A comparison problem may also develop if the RIA for the hormone is via a commercial kit versus a laboratory developed technique. In such comparisons, absolute numbers are difficult to contrast directly to one another and the reader must look at relative changes for the most part. The RIA technique is a relatively easy bioassay to use, as well as inexpensive, and has a high degree of reliability, but there is a limitation: the results represent the immunological activity of the hormone and not necessarily the biological activity. The biological activity depends not only on hormonal levels but also on receptor availability and sensitivity within the individual subject.
Finally, examination of the research protocols of studies that compare the hormonal status of exercising and non-exercising subjects reveals several problems. For example, to examine the effects of endurance training most studies have compared aerobically trained men with untrained sedentary-control men, in a retrospective fashion. The problem with this approach is the trained men have subjected themselves to the rigors of the training program, typically for years and the investigator is observing the end result. Because of the wide variations in response to training, as well as variations in the exact type of training, considerable divergence can be observed in the testosterone responses of these subjects. Another problem develops from the large discrepancy in what individual authors define as exercise training and exercise-trained states. For example, some researchers may consider an individual as chronically trained if they have performed regular exercise for one year, while other research may indicate that a five-year period is necessary. Additionally, one of the principal assumptions made within these retrospective studies is that the observed effects on testosterone levels are the results of the exercise training performed by the subject. Obviously, this assumption is not valid unless such things as psychological stresses, sleep loss, diet, weight loss, and hereditary factors, all of which affect testosterone level, are controlled.
Changes at Rest
The results of the retrospective comparative studies examining isolated, single blood samples suggest lower testosterone levels in chronically endurance-trained males. The subjects in these studies have typically been distance runners who had been involved with the physical training aspects of their sport for 1 to 15 years. In these studies, testosterone levels of the endurance-trained men were found to be 60-85% of the levels of matched, untrained men.
Prospective studies have also been conducted in which blood samples have been collected over days or weeks while exposing subjects to endurance training regimens. Findings thus far have been inconsistent, as some reports reveal significant reductions in resting testosterone following 1 to 6 months of intensive training, while others found no significant resting testosterone changes after 2 to 3 months of training. Differences in the initial training status of the subjects or the training administered may explain these discrepancies.
Endurance-trained males with lower testosterone also display other reproductive hormonal abnormalities. The most frequently reported changes involve decreased resting levels of prolactin and more importantly, no significant elevations in resting LH, even though there is decreased testosterone. These findings of altered prolactin and LH levels at rest in endurance trained men with low testosterone have been labeled by some researchers as a dysfunction of the hypothalamic-pituitary-testicular (HPT) regulatory axis. These findings with prolactin and LH have been reported in retrospective and prospective studies.
Several investigators have also conducted retrospective investigations where resting blood samples were collected every 20 or 30 min for 4- to 8-hour periods from chronically endurance-trained men and untrained controls. Results are similar to those of the isolated-sampling studies: resting testosterone concentration of the trained subjects were typically 70-80% of those found in the controls. As with isolated blood sampling studies, resting LH levels have not been significantly elevated. Again, these findings have been cited as a dysfunction in the HPT axis. These studies have been performed mainly with endurance runners, but it is likely that the effects apply to endurance athletes generally.
Mechanisms of HPT Axis Dysfunction. Several studies have attempted to elucidate the mechanism of the proposed HPT axis dysfunction found in endurance-trained men. These studies have focused on examining whether the dysfunction is central (hypothalamic or pituitary) or peripheral (testicular).
Centrally, the research work has focused upon alterations in LH release and/or prolactin release. Alterations in LH and prolactin release have been an area of extensive research relative to the study of exercising women who develop reproductive dysfunction. An exaggerated prolactin release to an exogenous stimulus of drugs or synthetic hormones has been found in endurance-trained males with low testosterone. Conversely, an attenuated release of LH due to doses of GnRH has been detected in endurance-trained males. Prolactin presents an interesting paradox in reproductive physiological function. Small amounts of prolactin seem necessary to work synergistically at the testicle with LH, while excessive levels disrupt both central and peripheral aspects of the HPT axis. Several investigators have found this altered prolactin and LH release. Figures 1 illustrates the findings of Hackney and associates for this unusual prolactin and LH hormonal response in trained males. Interestingly, although overall LH response is decreased, results are contradictory as to whether LH pulsatile characteristics (i.e., pulse frequency and amplitude) are affected by endurance training.
|
|
|
|
Peripherally, alterations in the sensitivity of the testes to LH changes has been the key research focus. Evaluations of testicular sensitivity to an exogenous stimulus have shown trained and untrained males to have comparable responses. Thus, these overall findings would suggest the development of a central problem in the HPT axis; however, data in this area are limited and not yet definitive.
An acute exercise bout could induce transient increases in circulating prolactin and cortisol, which could account for the observed changes in testosterone. The likelihood, however, of either of these transient states being a causative factor is doubtful. These are relatively small, temporary changes in prolactin and cortisol levels in response to exercise. Generally, the lowered testosterone associated with increased prolactin or cortisol is due to chronic supra-physiological elevations of these hormones that usually result from pathological states such as pituitary or adrenal tumors.
Changes in Response to Exercise
It is generally accepted that short-term maximal exercise results in an elevation of circulation testosterone levels. It is still an issue of some debate whether this effect is due simply to hemoconcentration effects, reduced metabolic clearance and/or possibly a hormone-mediated increase in testicular production. Surprisingly few studies have performed direct comparisons of exercise testosterone responses between endurance-trained (with low testosterone levels) and untrained control men. There appear to be no major differences in response between the two types of subject, but this area is in need of further investigation.
Typically testosterone responses to submaximal exercise are somewhat variable and seem dependent on the duration and intensity of exercise. A progressive increase in testosterone occurs during moderate exercise lasting 45 to 90 minutes, but 90 minutes of submaximal exercise has also been reported to produce no change or slight decreases in testosterone concentration. Exercise of a moderate to hard intensity performed until exhaustion or of more than 2 hours duration results in lowered testosterone concentrations. These changes in testosterone during submaximal exercise may be due to the interaction of several factors. Initially with submaximal exercise, hemoconcentration may produce elevations in testosterone levels. However, as the exercise continues there are reductions in testicular production and secretion. These production/secretion reductions appear to be principally the effects of declining testicular blood flow. Concurrently with the reduced testicle blood flow, hepatic blood flow may also be compromised, resulting in a declining hepatic clearance, which may further elevate testosterone levels. This latter change may be offset by an increased uptake by skeletal muscle as exercise duration continues, resulting in gradual reduction in testosterone levels. The acute effects of maximal and submaximal exercise on testosterone levels last a relatively short period of time (~1 to 48 hours).
Again, few studies seem to have compared submaximal exercise testosterone responses between endurance-trained (with low testosterone levels) and untrained control men. There is no evidence to suggest extensive response differences between the two types of subjects.
Physiological Impact of Lower Testosterone
Currently, there is little evidence to suggest that the development of low resting testosterone levels in men doing endurance training has any major detrimental effect on the testosterone-dependent physiological processes of the body. There is the potential for disruption in any of the processes influenced by testosterone, and obviously reproductive capacity is of key concern. Few studies have systematically studied this issue and currently only a few reports of decreased spermatogenesis or oligospermatic conditions existing in endurance trained males have been published. Additionally, some investigators have reported endurance trained men may have a lowered sex drive; but a direct cause-and-effect linkage between a lower sex drive and testosterone did not exist in these data.
There are also apparently no significant detrimental effects of lower testosterone on the other anabolic-androgenic effects of testosterone (protein synthesis and muscle mass development). However, this area has not been examined thoroughly by the exercise science community. An additional area in need of research concerns bone demineralization and low testosterone. Clinically, there is strong evidence indicating hypogonadal and hypo-testosteronemic men have severe mineral loss from their bones. There are no conclusive findings of mineral content change in endurance trained men with low testosterone, but several compelling case studies have been reported in the literature.
On the other hand, there could be beneficial physiologic adaptations from the lowered testosterone. Some researchers indicate that lowering testosterone may have cardiovascular protective effects and decrease the risk of coronary heart disease. A recent study out of Germany has provided strong evidence supporting this claim. This research has demonstrated that chemically induced reductions in endogenous testosterone level results in significant increases in high-density lipoprotein in men. Whether the lowering of testosterone in men performing endurance training will produce the same effect remains to be determined.
Endurance exercise training would seem to have significant effects upon the major male reproductive hormone, testosterone. A growing body of evidence suggests at rest testosterone is lowered in the endurance-trained male. The mechanism of this lowering is currently unclear, but may be related to dysfunction within the HPT regulatory axis brought about by months or years of endurance training. Currently, the time course of the changes in the reproductive endocrine system are unresolved. There are only limited findings to indicate that endurance training disrupts testosterone-dependent anabolic or androgenic processes in the male. The alterations in testosterone levels brought by endurance exercise training could even have cardiovascular protective effects.
Acknowledgment
The author is grateful for the help of H. Herbert Allred in the preparation of this manuscript.
Arce, J.C., & DeSouza, M.J. (1993). Exercise and male factor infertility. Sports Medicine, 15, 146-169.
Ayers J.W.T., Komesu, V., Romani, T., & Ansbacher, R. (1985). Anthropomorphic, hormonal, and psychologic correlates of semen quality in endurance-trained male athletes. Fertility Sterility, 43, 197-921.
Behre, H., Kliesch, S., Leifke, E., Link, T., Nieschlag (1997). Long-term effect of testosterone therapy on bone mineral density in hypogonadal men. Journal of Clinical Endocrinology and Metabolism, 82, 2386-2390.
Blair S.N., Kampert, J.B., Kohl, H., Barlow, C., Macera, C., Paffenbarger, R.S. (1996). Influence of cardiorespiratory fitness and other precursors on cardiovascular disease and all-cause mortality in men and women. Journal of the American Medical Association, 276, 205-210.
Bunt J., Bahr, J.M., Bemben, D.A. (1987). Comparison of estradiol and testosterone levels during and immediately following prolonged exercise in moderately active and trained males and females. Endocrine Research, 13, 157-172.
Galbo H. (1983). Hormonal and metabolic adaptations to exercise. New York, New York: Thieme-Stratton Inc.
Hackney, A.C. (1989). Endurance training and testosterone levels. Sports Medicine, 8, 117-127.
Hackney, A.C. (1996). The male reproductive system and endurance exercise. Medicine and Science in Sports and Exercise, 28, 180-189.
Hackney, A.C., Dolny, D.G., Ness, R.J. (1988). Comparison of resting reproductive hormonal profiles in select athletic groups. Biological Sport, 4, 200-204.
Hackney, A.C., Sinning, W.E., Bruot, B.C. (1988). Reproductive hormonal profiles of endurance-trained and untrained males. Medicine & Science in Sports & Exercise, 20, 60-65.
Hackney, A.C., Sinning, W.E., Bruot, B.C. (1990). Hypothalamic-pituitary-testicular axis function of endurance-trained and untrained males. International Journal of Sports Medicine, 34, 949-954.
MacConnie, S.E., Barkan, A., Lampman, R.M., Schork, M., Beitins, I.Z. (1986). Decreased hypothalamic gonadotrophin-releasing hormone secretion in male marathon runners. New England Journal of Medicine, 315, 411-416.
von Eckardstein, A., Kliesch, S., Nieschlag, E., Chirazi, A., Assmann, G., Behre, H. (1997). Suppresion of endogenous testosterone in young men increases levels of high density lipoprotein subclass lipoprotein A-I and liporotein(a). Journal of Clinical Endocrinology and Metabolism. 82, 3367-3372.
Wheeler, G.D., Wall, S.R., Belcastro,
A.N., Cumming, D.C. (1984). Reduced serum testosterone and prolactin
levels in male distance runners. Journal of the American Medical
Association, 252, 514-516.
newseditor=AT=sportsci.org · webmaster=AT=sportsci.org · Copyright
©1998
Edited by Mary Ann Wallace and Will
Hopkins. Webmastered by Jason Nugent
Last updated 13 Nov 98
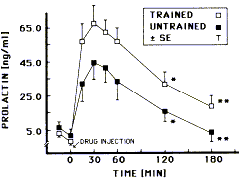 Figure
1A: Prolactin
release following an injection of a prolactin releasing
stimulant (metoclopramdide hydrochloride) in endurance
trained and untrained males. (Used with permission: Hackney
et al., 1990.)
Figure
1A: Prolactin
release following an injection of a prolactin releasing
stimulant (metoclopramdide hydrochloride) in endurance
trained and untrained males. (Used with permission: Hackney
et al., 1990.)
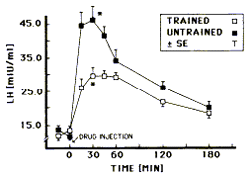 Figure
1B: LH release
following an injection of an LH-releasing stimulant
(gonadorelin hydrochloride) in endurance-trained and
untrained males. (Used with permission: Hackney et al.,
1990.)
Figure
1B: LH release
following an injection of an LH-releasing stimulant
(gonadorelin hydrochloride) in endurance-trained and
untrained males. (Used with permission: Hackney et al.,
1990.)